大肠杆菌噬菌体Bp4抗性菌株与其敏感菌株培养特性及耐药性检测(二)
2.3敏感菌株与抗性菌株基因组差异的分析
将敏感菌株与抗性菌株分别测序。结果显示,O78型敏感菌株与O78-R抗性菌株基因组全长均为4 798 435 bp。两组测序数据的比对分析结果显示,抗性菌株中有3个碱基发生突变,分别位于3个重叠群(contig)中(表1)。
表1显示,3个碱基的突变涉及到3个氨基酸,分别为乙醛脱氢酶的N→Y,十一碳二烯磷酸α-N-乙酰氨基葡萄糖磷酸转移酶1的W发生无义突变,以及UpfB蛋白L发生同义突变。致噬菌体发生抗性的原因很可能为contig00024中65 018 nt处的碱基突变,密码子由TGG变为TAG,编码的氨基酸由色氨酸突变为终止密码子,从而致蛋白质翻译提前终止,致使抗性菌株的葡萄糖磷酸转移酶由敏感菌株的372个氨基酸缩短为133个氨基酸,不能执行正常的酶功能(图3)。
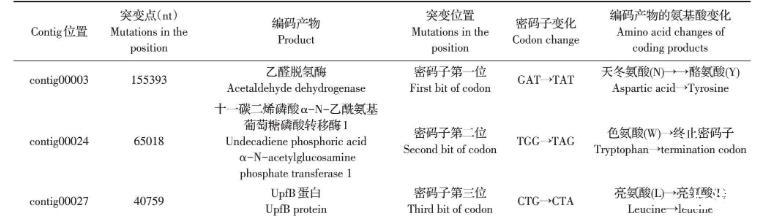
表1抗性菌株突变点的信息

图3敏感菌株O78与抗性菌株O78-R氨基酸序列比对
2.4敏感菌株与抗性菌株的血清型鉴定
采用玻板凝集法对O78型敏感菌株及O78-R抗性菌株进行血清型鉴定,结果显示,O78型敏感菌株与O78抗血清呈明显的凝集现象,液滴边缘出现麸皮样颗粒状的凝集片,液体清亮;O78-R抗性菌株与O78抗血清的凝集现象不明显,液体浑浊,且有少量细微颗粒。表明敏感菌株确为O78血清型,而抗性菌株血清型发生了改变,已不能鉴定为O78血清型。
2.5敏感菌株与抗性菌株的药敏试验
采用药敏纸片法分别检测O78型敏感菌株与O78-R抗性菌株的药物敏感性。结果显示,与敏感菌株相比,O78-R抗性菌株对替米考星、环丙沙星、氨曲南、妥布霉素的抑菌圈直径增大,耐药性降低(表2),表明噬菌体抗性菌株比其敏感菌株对以上抗生素更敏感。
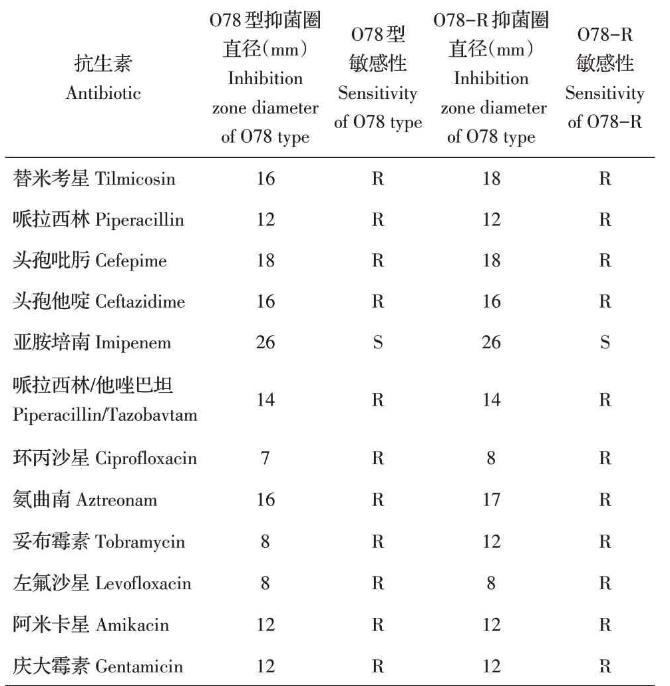
表2药敏试验结果
2.6敏感菌株与抗性菌株对小鼠的致病性试验结果
将敏感菌株与抗性菌株的增殖液经腹腔接种小鼠,记录小鼠的体况计分及存活率。结果显示,抗性菌株接种的小鼠未出现死亡,在感染后0~48 h小鼠体况计分一直维持在45分以上,之后略有下降(图4A);而敏感菌株接种的小鼠在接种后72 h内共死亡4只,体况计分比抗性菌株接种的小鼠约低30分(图4B),表明噬菌体抗性菌株对小鼠的致病性降低。

图4两组小鼠的体况计分(A)及存活率(B)统计结果
3讨论
为了研究噬菌体的抗性突变大肠杆菌菌株与其敏感菌株的差异,本研究经双层平板法筛选出对噬菌体具有抗性的大肠杆菌突变菌株,虽然在形态及培养特性上该抗性菌株与其敏感菌株未表现出差异,但该抗性菌株不再与O78抗血清发生强阳性的凝集反应,不再属于O78血清型。
基因组测序及分析结果显示,敏感菌株与抗性菌株基因组的全长一致,证明抗性菌株并无基因缺失和插入,但在抗性菌株中出现3处碱基的突变,导致1个同义突变,1个氨基酸替换(乙醛脱氢酶)以及1个无义突变。乙醛脱氢酶负责催化乙醛氧化为乙酸的反应,参与细菌的代谢,与噬菌体的感染无关。无义突变导致抗性菌株的十一碳二烯磷酸α-N-乙酰氨基葡萄糖磷酸转移酶1的翻译提前终止,而该蛋白参与细菌外膜中脂多糖O抗原的生物合成途径[10],导致脂多糖O抗原的合成受到抑制,提示该蛋白的突变是导致噬菌体不能感染抗性菌株的原因。本研究所用的噬菌体Bp4属于短尾噬菌体科,为N4类噬菌体[11],短尾噬菌体中N4类噬菌体G7C识别大肠杆菌表面的脂多糖受体[12],T7类噬菌体尾部蛋白识别的也是宿主菌表面的脂多糖受体[13],推测脂多糖也是噬菌体Bp4的受体。大肠杆菌为革兰氏阴性菌,细胞壁外膜中镶嵌大量的脂多糖[14]。革兰氏阴性菌的脂多糖由3部分组成:从内向外依次为脂质A、核心多糖、多糖侧链,多糖侧链又称为O-抗原,当脂多糖中的任何一部分缺失均会对脂多糖的合成造成影响。本实验中筛选的抗性菌株是由于碱基突变导致了色氨酸变为终止密码子,致脂多糖中参与O抗原生物合成的N-乙酰葡萄糖胺磷酸转移酶的翻译提前终止。通过凝集试验鉴定敏感菌株与抗性菌株的O抗原,结果显示敏感菌株为典型的O78血清型,而抗性菌株不再与O78抗血清发生凝集,不能鉴定为O78血清型。该结果证实O抗原是噬菌体Bp4识别并吸附宿主菌的关键受体。大肠杆菌的O抗原类型与其致病性有直接相关性,每株大肠杆菌只含有一种O抗原,能致鸡大肠杆菌病的常见血清型为O1、O2、O35及O78[15]。本研究获得的噬菌体抗性菌不再具有O78抗血清的凝集性,提示细菌的致病性可能下降。乐率等通过对铜绿假单胞菌PA1株与噬菌体的相互作用,筛选出抗性菌株PA1r并做了相关研究,发现抗性菌株可以分泌红色素,且致病性下降[6]。噬菌体抗性菌株致病性的下降已被多项研究所证实[4,16],本研究经腹腔注射敏感菌株小鼠的死亡率达到40%,而腹腔注射抗性菌株小鼠的死亡率为0,也证明了噬菌体抗性菌株致病性的下降。噬菌体抗性菌株致病性的下降会导致细菌在动物体内更容易被免疫系统清除。
本研究发现,噬菌体抗性菌株的耐药性发生了改变,其对替米考星、环丙沙星、氨曲南、妥布霉素的抑菌圈直径增大,比噬菌体敏感菌株的耐药性降低,该结果与Jalasvuori报道的结果类似[17]。噬菌体抗性菌株耐药性的下降会导致其在较低的抗生素浓度下即可被杀死。
噬菌体抗性菌株产生的几率大约为千万分之一。细菌对噬菌体的抗性是相对的,其对一株噬菌体产生抗性,但还可以被其他的噬菌体感染。于玲等分离到对大肠杆菌具有抗性的噬菌体,并将其与原噬菌体混合制成新的噬菌体鸡尾酒用于杀菌,结果表明,该噬菌体鸡尾酒不仅具有明显的杀菌效果,而且在体外降低了噬菌体抗性菌株的产生和突变频率[18]。
噬菌体抗性突变菌株的产生是细菌和噬菌体相互作用的结果,抗性菌株的产生必然会导致细菌基因组及表型的变化。本研究通过分析大肠杆菌短尾噬菌体Bp4的抗性菌株和敏感菌株,发现抗性菌株的耐药性和致病性均下降,说明噬菌体抗性菌株的产生不会对噬菌体治疗效果产生较多副作用,也为噬菌体治疗中出现抗性菌问题的解决提供了参考依据。
大肠杆菌噬菌体Bp4抗性菌株与其敏感菌株培养特性及耐药性检测(一)
大肠杆菌噬菌体Bp4抗性菌株与其敏感菌株培养特性及耐药性检测(二)
相关新闻推荐
1、综述微生物修复菲污染中降解菌的菌属、降解机理、分子机制、影响因素(一)
2、李斯特菌ST87在25°C不同营养条件下牛肉和蔬菜汁中的生长曲线