短梗霉素A对植物病原真菌灰葡萄孢菌AUR 1基因的抑制生长机理(二)
2、结果与分析
2.1 AbA敏感性测定
结果表明,野生株Bc AUR1在2μg/m L Ab A的培养基上能够生长;Ab A为8μg/m L时,生长受到严重抑制;当Ab A浓度达到50μg/m L时,未见菌体生长(图2)。突变株Bc AUR1a在Ab A浓度为2~50μg/m L的培养基上均能生长(图2),表明突变菌株Bc AUR1a对Ab A产生了抗性。

图2灰葡萄孢菌对AbA敏感性测定
2.2突变株Bc AUR1a对Ab A处理柑橘的侵染
2.5%甲醇处理的对照组柑橘全部被野生株Bc AUR1感染(图3a),50μg/m L Ab A处理的试验组柑橘均未被野生株Bc AUR1感染(图3b),说明Ab A抑制了野生株Bc AUR1对柑橘的侵染,能够有效防治柑橘灰霉病的发生。而突变株Bc AUR1a则对试验组和对照组柑橘都能侵染(图3c~d),说明Ab A不能抑制突变株BCAUR1a对柑橘的侵染,突变株Bc AUR1a产生了Ab A抗性。
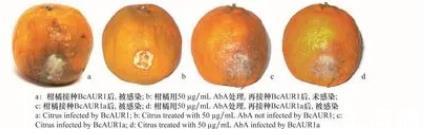
图3 AbA对灰葡萄孢菌侵染柑橘的影响
2.3 Ab A对灰葡萄孢细胞形态的影响
未经Ab A处理的灰葡萄孢Bc AUR1细胞壁较厚,细胞壁表面界限和表面物质清晰可见;细胞质膜平滑连续,紧贴细胞壁;内膜系统丰富,线粒体膜完整,嵴丰富而粗壮,液泡膜完整,细胞质中富含糖原颗粒(图4a~b)。在8μg/m L的Ab A作用下,细胞明显变小,细胞壁变薄,细胞壁和细胞质膜表面粗糙不平,皱缩明显,部分质膜和液泡膜破裂;内膜系统的部分膜界限不清晰,线粒体肿胀,糖原颗粒和细胞内容物显著减少(图4c~f)。表明Ab A抑制了IPC合成酶的活性,导致鞘脂类合成代谢受阻,细胞膜结构受到严重影响,质膜和内膜系统形态异常,发生褶皱甚至破裂,造成细胞内糖原和内容物泄漏。
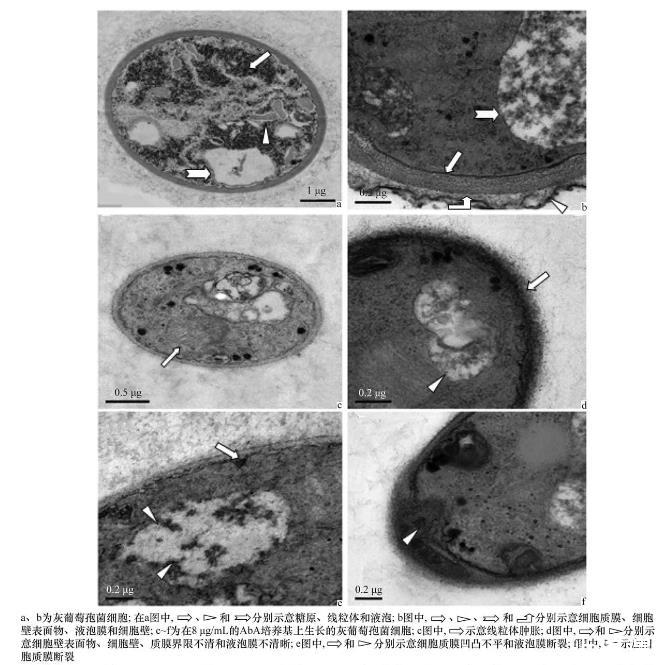
图4灰葡萄孢菌细胞形态的透射电镜
3、讨论
Ab A能抑制多种真菌的生长,对假丝酵母(Candida)、曲霉(Aspergillus)、酿酒酵母(Saccharomyces cerevisiae)等病原真菌均有抑制作用。我们通过Ab A对灰葡萄孢菌野生株BcAUR1及其AUR1基因但内含子缺失的突变株Bc AUR1a抑制作用的研究,也证实低浓度Ab A(8μg/m L)能够显著抑制灰葡萄孢菌野生株Bc AUR1的生长,高浓度Ab A(50μg/mL)则完全抑制其生长,但突变株Bc AUR1a则不受Ab A的影响,在高浓度Ab A存在下亦能正常生长。灰葡萄孢菌感染柑橘的试验也证实了这一结果。这些结果均表明Ab A通过抑制IPC合成酶的活性,导致鞘脂合成不足,从而抑制了灰葡萄孢菌的生长。试验中突变株BcAUR1a是AUR1基因中缺失内含子,产生了AbA抗性,抵抗Ab A对灰葡萄孢菌生长的抑制,说明该内含子可能在AUR1基因表达调控中发挥重要的作用,目前已有许多研究表明内含子具有调控基因转录的功能,在其序列中发现了转录调控元件,起着启动子、增强子和抑制子的作用。
鞘磷脂是细胞膜的重要组成成分,特别是生物膜脂筏(lipid raft)结构中的主要物质,鞘磷脂还与细胞信号转导、胞吞、胞饮等密切相关。本研究发现,Ab A导致灰葡萄孢细胞质膜和内膜系统形态异常,质膜和液泡膜断裂,导致细胞内物质泄露,从而抑制了细胞生长。进一步证实了Ab A使神经酰胺和磷脂酰肌醇生成肌醇磷脂酰神经酰胺的反应受阻,导致鞘磷脂类物质的合成量不足,细胞膜结构遭到破坏,造成糖原等细胞内物质泄露。
综上所述,Ab A抑制丝状真菌生长的机理是抑制了IPC合成酶的活性,导致鞘磷脂类物质的合成不足,细胞膜结构遭到破坏,细胞内物质外漏。
短梗霉素A对植物病原真菌灰葡萄孢菌AUR 1基因的抑制生长机理(一)
短梗霉素A对植物病原真菌灰葡萄孢菌AUR 1基因的抑制生长机理(二)