4株金黄色葡萄球菌噬菌体形态、生物学特性、生长曲线及基因组特征(三)
2.6一步生长曲线
将噬菌体SAPYZU-04、SAPYZU-15、SAPYZU-SapM13和SAPYZU-SapM14分别与宿主菌混合后培养,得到噬菌体的一步生长曲线。如图6所示,烈性噬菌体SAPYZU-04和SAPYZU-15侵染宿主的10 min内效价无明显变化,表明它们的潜伏期均为10 min。在10~150 min内,噬菌体SAPYZU-04和SAPYZU-15的效价一直保持升高趋势,随后趋于稳定,表明它们的裂解期均约为140 min,裂解量分别约为每个细胞210和322 PFU。而温和噬菌体SAPYZU-SapM13和SAPYZU-SapM14的潜伏期较长,分别约为15和30 min;裂解期较短,分别约为30和45 min,裂解量分别为每个细胞52和49 PFU。上述结果显示,烈性噬菌体SAPYZU-04和SAPYZU-15的裂解量均高于Gutierrez等分离的金葡菌烈性噬菌体(每个细胞15和25 PFU)。温和噬菌体SAPYZU-SapM13和SAPYZU-SapM14的裂解量比Feng等分离的温和噬菌体JD419裂解量(每个细胞33 PFU)更高。潜伏期和爆发量是表示噬菌体裂解能力的重要生物学特性,具有较短潜伏期和较大爆发量的噬菌体更适合作为生物防治剂。
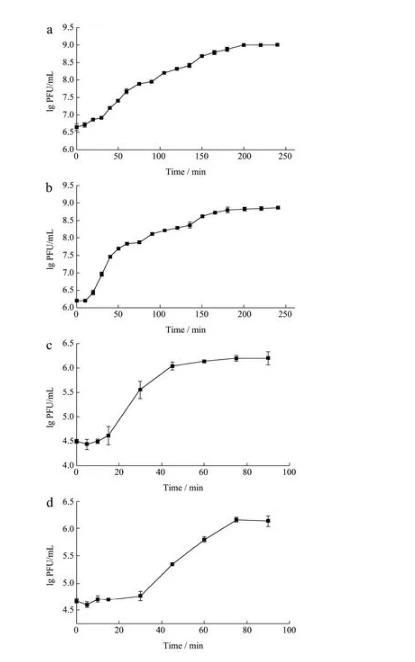
图6噬菌体的一步生长曲线
2.7系统发育树
全基因组序列分析结果显示烈性噬菌体SAPYZU-04和SAPYZU-15基因组全长分别为140 584 bp和135 178 bp,而温和噬菌体SAPYZU-SapM13和SAPYZU-SapM14的基因组全长分别为44 922 bp和44 232 bp。其中噬菌体SAPYZU-04的GC含量为31.48%,预测到188个ORFs;噬菌体SAPYZU-15的GC含量为29.90%,预测到187个ORFs;噬菌体SAPYZU-SapM13的GC含量为33.98%,包含71个ORFs;噬菌体SAPYZU-SapM14的GC含量为33.89%,包括73个ORFs。
截至2021年4月,GenBank数据库中记录192个金黄色葡萄球菌噬菌体基因组序列。本研究通过这些金黄色葡萄球菌噬菌体基因组序列进行系统发育分析,解析4株噬菌体和其他金黄色葡萄球菌噬菌体之间的关系。此树形图(图7)划分了3个主要的遗传谱系(I-Ⅲ)。聚类分析结果表明,烈性噬菌体SAPYZU-04和SAPYZU-15与Hellelleviridae科的噬菌体聚在一个谱系中(谱系Ⅱ),且SAPYZU-04和SAPYZU-15与噬菌体qdsa001基因组核苷酸序列最相似,相似性>99.0%;温和噬菌体SAPYZU-SapM13和SAPYZU-SapM14与Azeredovirinae亚科Dubowvirus属的噬菌体聚在一个谱系中(谱系Ⅲ),且SAPYZU-SapM13和SAPYZU-SapM14与TEM126和SAP26的基因组核苷酸序列一致性>95%。因此,根据ICTV2022年最新的分类标准,噬菌体SAPYZU-04和SAPYZU-15属于Hellelleviridae科;而SAPYZU-SapM13和SAPYZU-SapM14属于Azeredovirinae亚科Dubowvirus属。
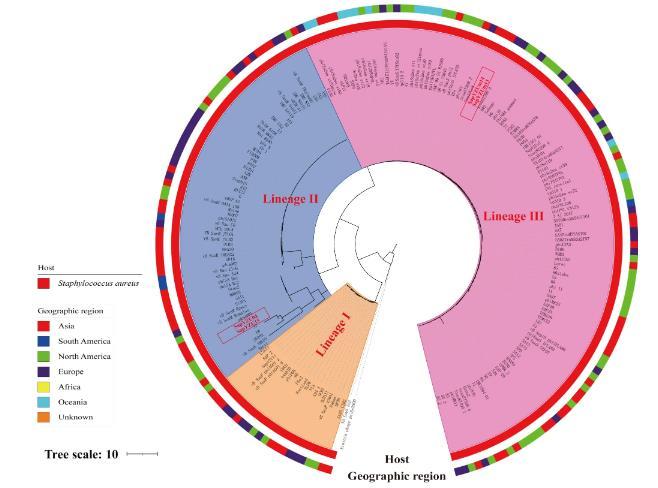
图7系统发育分析
2.8全基因组序列分析
对4株噬菌体基因组携带基因的蛋白功能进行注释,结果如图8所示。噬菌体SAPYZU-04的基因组编码32个DNA代谢相关基因,包括核酸内切酶基因(orf38)、DNA解旋酶基因(orf115)和DNA聚合酶Ⅰ基因(orf144)等;噬菌体SAPYZU-15的基因组包含多个DNA代谢基因,其中包括3个DNA合成相关基因(orf1、orf3和orf5)、2个DNA聚合酶Ⅰ(orf10、orf11)和DNA绑定蛋白(orf82);噬菌体SAPYZU-SapM13的基因组编码13个与噬菌体DNA复制、调节和转录相关的基因,包括核酸外切酶基因(orf7)、转录因子(orf9)、DNA复制基因(orf25)和DNA解旋酶(orf45)等;而噬菌体SAPYZU-SapM14编码11个与噬菌体DNA代谢相关的基因,包括转录因子(orf11)、DNA绑定蛋白(orf24)、DNA解旋酶(orf28)和核酸内切酶(orf51)等。其中SAPYZU-15基因组编码丰富的DNA合成基因加速了噬菌体大分子的合成。SAPYZU-04的基因组编码3个DNA组装基因(orf66、orf68和orf69),SAPYZU-15的基因组编码1个DNA组装基因(orf117),SAPYZU-SapM13和SAPYZU-SapM14均包含2个DNA包装基因(分别为orf48、orf49、orf52和orf53)。其中SAPYZU-15编码的DNA包装基因与噬菌体LSA2308的氨基酸序列具有95.9%的相似性;LSA2308的爆发量约为407 PFU/cell。DNA包装蛋白可以有效地将DNA泵入尾部噬菌体原衣壳中,加速噬菌体的组装。噬菌体SAPYZU-04和SAPYZU-15的基因组中分别包含3个和1个尾部蛋白基因(分别为orf108、orf110、orf158和orf142),噬菌体SAPYZU-SapM13编码5个尾部相关蛋白基因,包括尾管蛋白基因(orf57)和主要尾部蛋白基因(orf58和orf62)等;而SAPYZU-SapM14编码6个尾部相关蛋白基因,包括尾部蛋白基因(orf57、orf61、orf62)等。这些独特的尾部蛋白基因可能是此4株噬菌体具有广泛的裂解谱的原因。其中SapYZU-15含有的尾蛋白基因与噬菌体KSAP11的基因组氨基酸序列有95.90%的相似性。KSAP11具有广泛的裂解谱,对30株SA菌株(包括MSSA和MRSA菌株)具有裂解活性。噬菌体SAPYZU-04包含1个穿孔素基因(orf53)和3个裂解酶基因(orf52、orf100和orf102),SAPYZU-15包含1个穿孔素基因(orf107)、5个裂解酶基因(orf103、orf105、orf149、orf153和orf155);而SAPYZU-SapM13和SAPYZU-SapM14的基因组均只包含1个穿孔素基因(分别为orf70和orf2)和1个裂解酶基因(分别为orf71和orf3)。穿孔素基因在裂解过程中诱导宿主细胞膜去极化,调节裂解时间。其中SAPYZU-15基因组中有2个裂解酶基因(orf103和orf155)与MR003具有较高的氨基酸序列相似性(97.90%和99.00%),MR003是高效的SA烈性噬菌体。因此,在这些基因的协同作用下,噬菌体SAPYZU-04和SAPYZU-15潜伏期更短,裂解量更高。此外,温和噬菌体基因组中的整合酶基因和CI阻遏蛋白是决定其生命周期的重要因素,整合酶基因通过催化两个DNA分子之间重组使得噬菌体在宿主染色体中进行基因的整合和切除,并且温和噬菌体通过CI阻遏蛋白阻断裂解基因的转录和翻译,从而进入溶原周期;而抗阻遏蛋白控制噬菌体在侵染时进入裂解周期。本文的温和噬菌体SAPYZU-SapM13和SAPYZU-SapM14基因组编码特异性整合酶证明它们具有溶原性,均预测到抗阻遏蛋白,但是未预测到CI阻遏蛋白和噬菌体抗性基因,可能导致温和噬菌体SAPYZU-SapM13和SAPYZU-SapM14在宿主谱测定时易进入裂解周期,上述结果与Tian等的研究结果一致。

图8全基因组分析
3、结论
金黄色葡萄球菌严重威胁食品安全,同时也是全球关注的重大公共卫生问题之一。因此,迫切需要开发安全可靠的生物防治方法,以提高公共卫生水平,确保消费者安全。本研究分离的烈性噬菌体SAPYZU-04和SAPYZU-15效价均达到109PFU/mL,而且具有较宽的裂解谱,两株噬菌体对51株SA菌株的裂解率均高达100%;在40~60℃温度范围内、pH值在3~12范围内均保持良好活性;潜伏期较短、裂解能力强,裂解量分别高达每个细胞210和322 PFU。全基因组序列分析结果显示SAPYZU-04和SAPYZU-15均包含多种DNA代谢基因、多个裂解酶基因、独特的DNA包装基因及尾部蛋白基因。因此,烈性噬菌体SAPYZU-04和SAPYZU-15是一种更合适、更有前景的生物防治选择,有望应用于控制食品中金葡菌的污染。
4株金黄色葡萄球菌噬菌体形态、生物学特性、生长曲线及基因组特征(一)
4株金黄色葡萄球菌噬菌体形态、生物学特性、生长曲线及基因组特征(二)
4株金黄色葡萄球菌噬菌体形态、生物学特性、生长曲线及基因组特征(三)
相关新闻推荐
1、微生物生长曲线监测系统记录单核细胞增生李斯特菌在不同处理条件下的生长过程
2、适用于低盐发酵香肠葡萄球菌与乳酸菌生长特性及产酸能力测定及筛选(二)
3、恒河猴肺中阴沟肠杆菌的分离鉴定及药物敏感性试验——摘要、材料和方法