棉子糖抑制变异链球菌生物膜形成机制研究进展(一)
变异链球菌(Streptococcus mutans)是一种具有代表性的生物被膜细菌,它通过葡糖基转移酶(glucosyltransferase,GTF)活性引起龋齿。葡聚糖是由GTFs从蔗糖中合成的,为S.mutans提供了紧密附着在牙釉质上的结合部位。因此,如果开发出一种干扰GTF功能的新化合物,就有可能控制S.mutans的生物被膜形成。我们发现,天然产物中的一种低聚糖棉子糖能强烈抑制生物膜的形成、GTF相关基因的表达和葡聚糖的产生。此外,通过减少细菌黏附对唾液包裹的羟基磷灰石(hydroxyapatite,HA)生物膜的抑制作用表明棉子糖在口腔健康中的适用性。棉子糖的这些作用似乎是由于其调节S.mutans GTF活性的能力所致。因此,棉子糖可以被认为是一种抗生物被膜剂,用作口腔用品和预防龋齿的牙科材料的物质。
主要内容
1.棉子糖对S.mutans生物被膜形成的影响
棉子糖减少了大多数从人类口腔分离的链球菌(KCOM 1136至KCOM 1228)的生物被膜形成(图1)。与未经处理的生物膜(即对照生物膜)相比,当添加100µM或1000µM棉子糖时,S.mutans和远缘链球菌(Streptococcus sobrinus)的生物膜形成平均分别被抑制12-25%和11-17%。
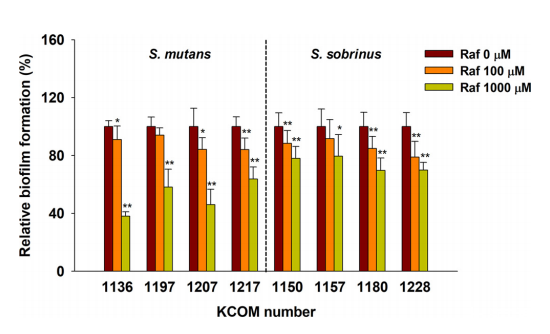
图1棉子糖处理24小时后的链球菌生物膜形成
在静态条件下,棉子糖处理(0~1000µM)后形成的S.mutans和S.sobrinus的生物膜
误差条表示五个测量值的标准偏差。**,P<0.005;*,P<0.05,与对照组比较。Raf,棉子糖,下同
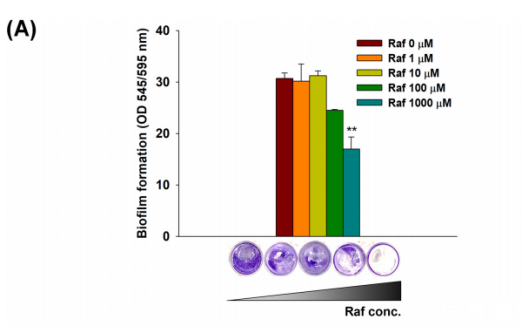
特别是,本研究中具有代表性的生物被膜菌S.mutans KCOM1136在棉子糖处理后生物被膜的形成显著减少,经1000µM棉子糖处理后生物被膜的形成比对照生物膜减少了50%以上。在静态和流动条件下分析了棉子糖处理对KCOM 1136生物膜形成的抑制作用。在静态条件下,当棉子糖浓度较高(1000µM)时,生物膜的形成显著减少44%(图2A)。在流动条件下,尽管对照生物膜和棉子糖处理生物膜的形态相似,但棉子糖处理生物膜的平均体积和厚度比对照生物膜减小了54%-64%(图2B)。
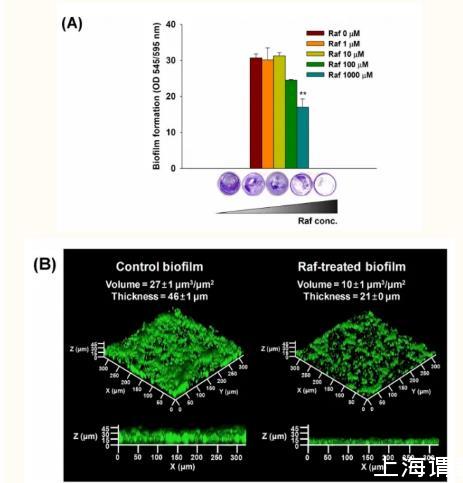
图2静态和流动条件下棉子糖处理后S.mutans生物膜的形成
(A)棉子糖处理(0-1000µM)后的CV染色生物膜;在静态条件下(OD545/OD595)测量OD进行定量。(B)基于激光共聚焦显微镜图像的DAPI染色生物膜的体积和厚度;在流动条件下,向S.mutans生物膜中加入1000µM棉子糖处理48h。
棉子糖是一种由半乳糖和蔗糖组成的三糖。然而,在蔗糖和半乳糖分别处理S.mutans生物膜,可以观察到这呈现相互矛盾的现象。半乳糖显示出类似于棉子糖的生物被膜抑制活性;相反,蔗糖促进了生物被膜的形成。此外,蔗糖在1000µM时促进S.mutans的生长,而棉子糖处理(0-1000µM)对S.mutans的生长没有影响。
2.棉子糖抑制S.mutans生物被膜的机制
棉子糖处理后,对S.mutans的蔗糖消耗进行了24 h的监测,以探讨棉子糖与蔗糖的关系。用来评估蔗糖消耗的方法适合于检测蔗糖浓度,但不能响应高浓度的棉子糖,如蔗糖和棉子糖的标准曲线所示(见图S3)。S.mutans在24小时内消耗了88%的蔗糖(图3A)。然而,随着棉子糖和蔗糖浓度的增加,S.mutans的蔗糖消耗量比对照(即不加棉子糖的处理)减少。当10μM、100μM和1000μM棉子糖与100μM蔗糖一起使用时,24 h内分别消耗79%、70%和56%的蔗糖。
图S3蔗糖和棉子糖的标准曲线(0-1000μM),用于评估蔗糖消耗试验的适用性
此外,将不同浓度的棉子糖和蔗糖(0-1000μM)同时加入到S.mutans的培养基中,分析了竞争生物膜的形成。与对照相比,1000μM棉子糖处理后生物被膜形成减少了50%,而同时处理1000μM蔗糖后生物被膜形成减少了26%(图3B,左)。这一发现表明,棉子糖处理后对生物膜形成的抑制随着蔗糖浓度的增加而降低。同样,随着棉子糖浓度的增加,蔗糖处理后的生物膜形成减少(图3B,右)。这些结果表明,棉子糖和蔗糖在S.mutans的蔗糖消耗和生物被膜形成方面存在竞争关系。
通过分子对接研究了棉子糖在S.mutans中作为葡聚糖酶抑制剂的可能性。图3C显示了棉子糖在S.mutans葡聚糖酶活性部位的最佳对接姿势。棉子糖的对接位置与葡萄糖酶的活性部位相匹配,并与Asp593形成氢键,Asp593被认为是受体糖定位的关键氨基酸残基,影响S.mutans葡糖酶的转糖基化专一性。然而,蔗糖的对接姿势仅位于亚基1,它由Arg475、Asp477、Glu515和Asp909组成,没有与Asp593相互作用,如补充材料中的图S4A所示。棉子糖和蔗糖中常见的葡萄糖和果糖部分与亚基1位的氨基酸残基密切接触。对于D-半乳糖,最好的对接姿势也只与亚基1的氨基酸残基密切接触,包括Asp475、Asp477、Asp909和Gln 960(见图S4B)。特别是,棉子糖半乳糖部分C-3和C-4位的羟基与Asp593以及亚基1位的氨基酸残基相互作用,这可能增强其与葡聚糖酶的结合亲和力。总体而言,对接研究表明棉子糖、蔗糖和半乳糖可能与葡聚糖酶的亚基1位点接触,总分分别为9.90、7.88和6.42。
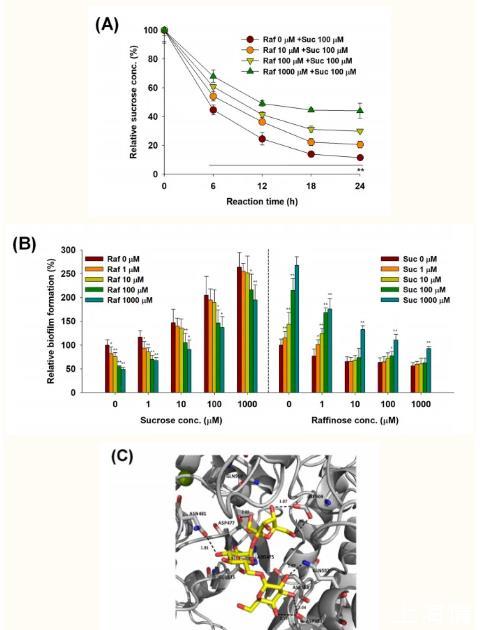
图3棉子糖处理后抑制S.mutans生物被膜形成的可能机制
(A)棉子糖处理的S.mutans生物膜细胞中的蔗糖消耗。在静态条件下,蔗糖(100μM)和棉子糖(0-1000μM)处理24小时后形成S.mutans生物膜。Suc,蔗糖。(B)棉子糖和蔗糖之间的竞争性生物被膜形成试验。S.mutans生物膜是在0-1000μM浓度的棉子糖和蔗糖处理后形成的。(C)S.mutans葡糖酶中棉子糖的最佳对接姿势)。
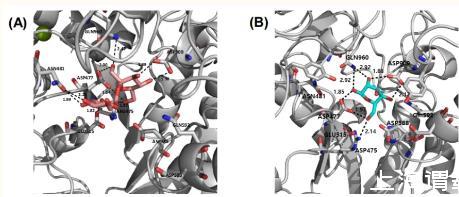
图S4(A)蔗糖和(B)D-半乳糖在S.mutans葡聚糖酶活性部位的最佳对接姿势。